A new perspective on how to slow aging
By Genomic Press
 |
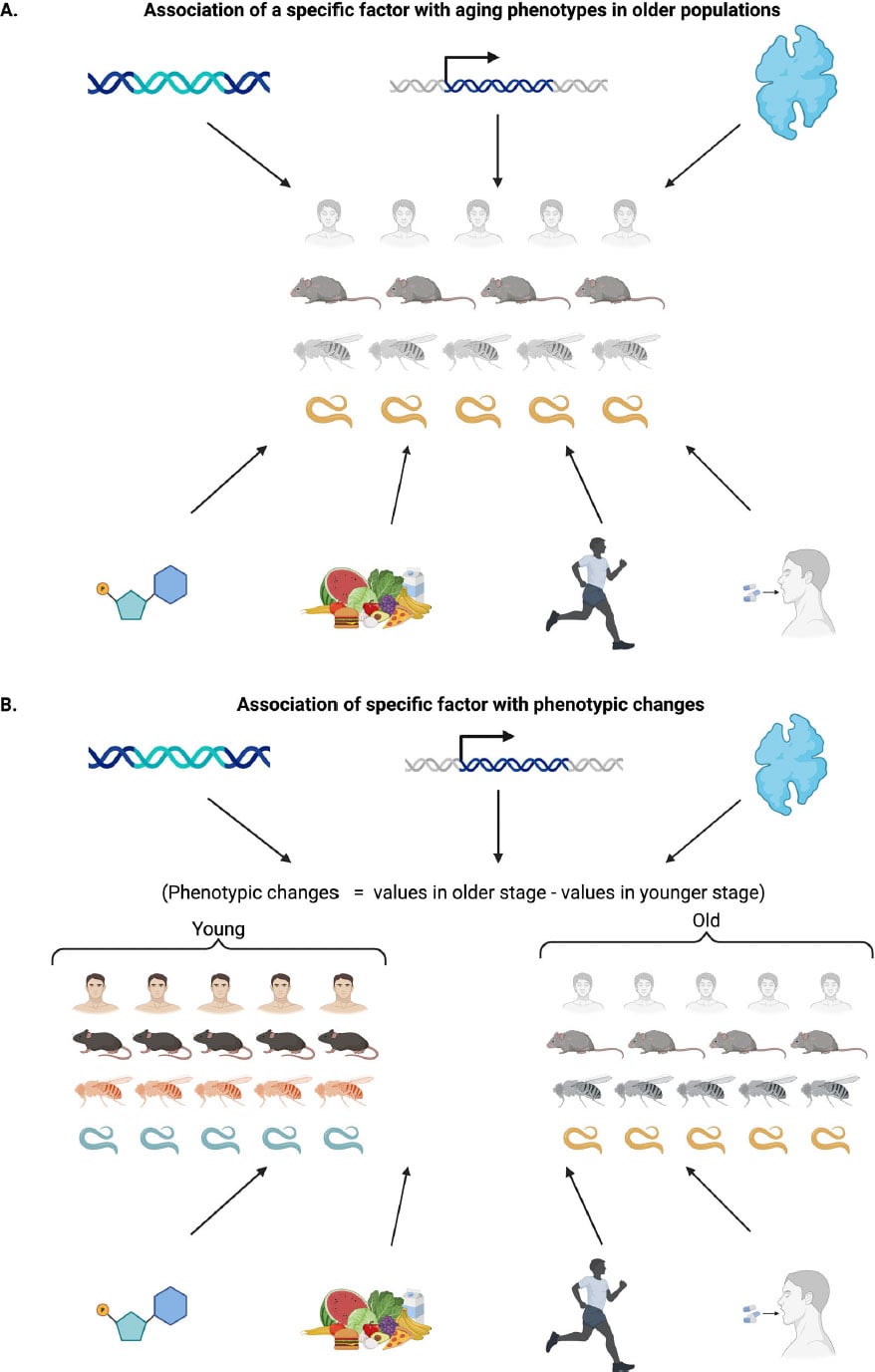
| Multidimensional nature of aging: phenotypic changes across levels of biological complexity. The figure illustrates time-dependent phenotypic change across molecular, cellular, tissue, and organismal scales in multiple species. Credit: Dan Ehninger |
The analysis, published in Genomic Psychiatry,
calls on researchers to rethink how biological aging is measured and
interpreted. Dr. Dan Ehninger, who leads the Translational Biogerontology
Laboratory at the German Center for Neurodegenerative Diseases, and Dr. Maryam
Keshavarz conducted a systematic review examining widely used indicators of
aging.
Their work argues that common measures such as lifespan
extension, epigenetic clocks, frailty indices, and even the widely cited
hallmarks of aging framework may blur the line between true changes in aging
and general physiological effects that occur regardless of age.
One of the most surprising insights comes from comparing
causes of death across species. In humans, cardiovascular disease is
responsible for roughly 35 to 70 percent of deaths among older adults. Autopsy
studies prove that even centenarians who appeared healthy shortly before death
almost always died from identifiable medical conditions rather than from old
age alone.
Research on people between 97 and 106 years old further supports this pattern, with vascular diseases remaining the leading cause of death. These findings highlight that even exceptional longevity usually ends with a specific disease.
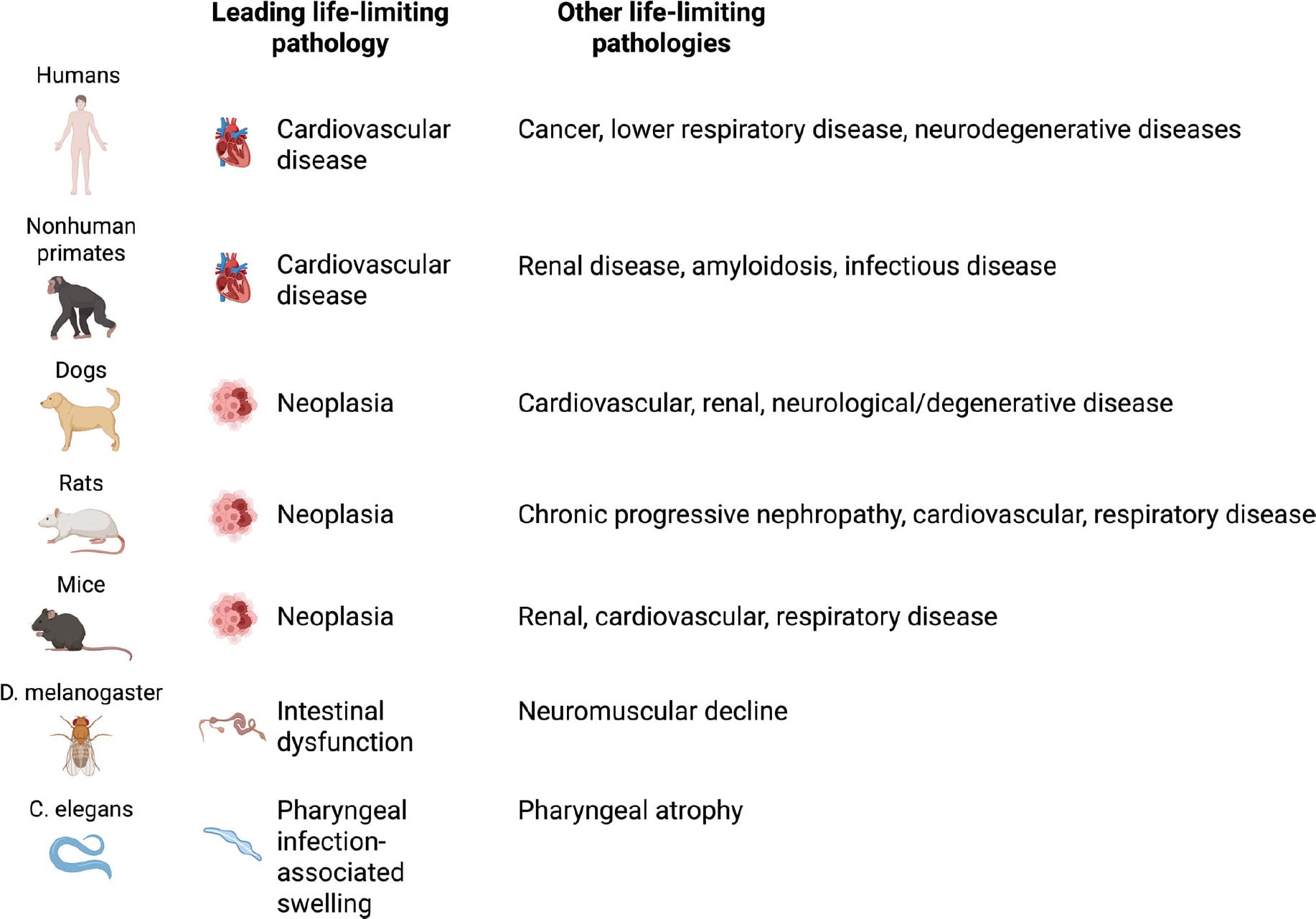
What Actually Causes Death in Aging Across Species
The main causes of death vary significantly between species.
In laboratory mice, cancer dominates and accounts for about 84 to 89 percent of
deaths associated with aging. Dogs show a similar trend, with nearly half of
older dogs dying from cancer. Among captive nonhuman primates, the pattern
looks closer to humans, as cardiovascular disease causes more than 60 percent
of deaths in aging rhesus macaques. Even small invertebrates display
species-specific limits on lifespan. Intestinal or neuromuscular failure often
limits survival in Drosophila, while pharyngeal infections and
deterioration drive mortality in C. elegans.
“This pattern illustrates that interventions targeting
specific pathologies can extend lifespan by addressing critical bottlenecks to
survival, but they do not necessarily slow the overall aging process,” the
authors write.
Understanding this distinction is important when considering
how human lifespan has increased over the past two centuries. In the past,
infectious diseases such as the bubonic plague, smallpox, and tuberculosis were
the main causes of death and killed millions of people. Advances in medicine
and public health, including vaccines, antibiotics, and sanitation
improvements, greatly reduced deaths from these diseases.
Lifespan Gains Do Not Always Mean Slower Aging
Yet according to the researchers, this historical shift
reflects changing causes of death rather than a slowdown in the biological pace
of aging. By reducing deaths from infectious disease, society delayed when
people die, but did not necessarily alter the underlying processes that drive
aging.
This observation has important implications for modern aging
research. If lifespan increases simply because one major disease is delayed,
then longer survival does not automatically mean that aging itself has slowed.
For example, an intervention that extends the lifespan of mice by delaying
cancer is fundamentally different from one that slows the gradual decline of
many body systems, even if both produce similar survival curves.
Epigenetic aging clocks, which estimate biological age
using DNA methylation patterns, have become widely used tools in
aging research. They can help classify individuals by risk level, detect
accelerated aging in populations, and monitor biological changes over time.
However, Ehninger and Keshavarz argue that these tools raise more profound
questions about what exactly they measure.
Limits of Epigenetic Aging Clocks
A major limitation is that aging clocks are based on
correlations. They identify molecular patterns associated with chronological
age, but those patterns may not directly cause aging. Instead, they may simply
reflect biological changes that happen as a result of aging.
The authors compare this to estimating a person’s age from
facial images. Wrinkles and gray hair can help predict age, but they reveal
little about the biological mechanisms that drive aging. Supporting this
concern, the review cites epigenome-wide Mendelian randomization studies
showing that traditional aging clocks are not strongly enriched for CpG sites
that play causal roles in aging.
Another issue is that most aging clocks capture a single
moment in time. If an intervention appears to reduce biological age,
researchers must determine whether aging has actually slowed or whether the
treatment simply shifted biomarker values. Even newer tools such as
DunedinPACE, which aim to estimate the pace of aging rather than static
biological age, often rely on biomarkers linked to age-related traits without
directly identifying underlying mechanisms.
Problems With Frailty Indices as Aging Measures
Frailty indices face similar challenges. These indices are
typically based on small sets of semiquantitative traits, such as fur
condition, spinal curvature, or the presence of tumors, scored using simple
categorical scales. As a result, they represent only limited aspects of
aging-related change.
Because these scores combine multiple traits into a single
number, each component is implicitly treated as equally important biologically.
Improvements in a single factor, such as reduced tumor burden, could lower the
overall frailty score and give the impression that aging has broadly improved,
even though the change reflects improvement in one specific disease.
The review also examines the hallmarks of aging framework,
first introduced in 2013 and expanded to twelve hallmarks in 2023. These
hallmarks include processes such as genomic instability, telomere shortening,
epigenetic alterations, disrupted protein maintenance, and cellular senescence.
The framework has strongly shaped research priorities and funding decisions.
The authors ask whether the available evidence truly supports the idea that
targeting these hallmarks slows aging.
Do the Hallmarks of Aging Truly Drive Aging?
To investigate this question, Keshavarz and Ehninger
analyzed the primary studies cited as evidence for each hallmark. Their review
focused on research used to claim causal relationships between these processes
and aging.
They found a major methodological limitation. Between 56.86
and 99.96 percent of the phenotypes used as supporting evidence were examined
only in older animals. Researchers often did not test the same interventions in
young treated groups. Without these comparisons, it is impossible to determine
whether treatments actually slow aging or simply create age-independent
baseline effects.
When studies did include young animals, interventions often
affected both age groups. Across the research cited for the hallmarks
framework, the authors identified 602 phenotypes tested in young animals. Of
these, 436, or 72.4 percent, showed similar effects in young groups. This
suggests that many observed changes reflect baseline physiological shifts
rather than true antiaging mechanisms.
“Consequently, the evidence cited for most hallmarks
supports the presence of general physiological effects rather than true
antiaging mechanisms,” the review concludes.
Distinguishing True Antiaging Effects From Baseline
Changes
The researchers propose a framework to better distinguish
genuine aging interventions from general physiological effects. They describe
three categories of treatment outcomes for age-sensitive phenotypes.
Rate effects occur when an intervention reduces the slope of
age-related change, which would indicate that it is modifying the processes
driving aging. Baseline effects occur when similar changes appear in both young
and old animals, suggesting the treatment works independently of aging. Mixed
effects occur when both groups change, but older animals show stronger
responses, which requires careful interpretation because it may reflect
combined mechanisms or differences in treatment duration.
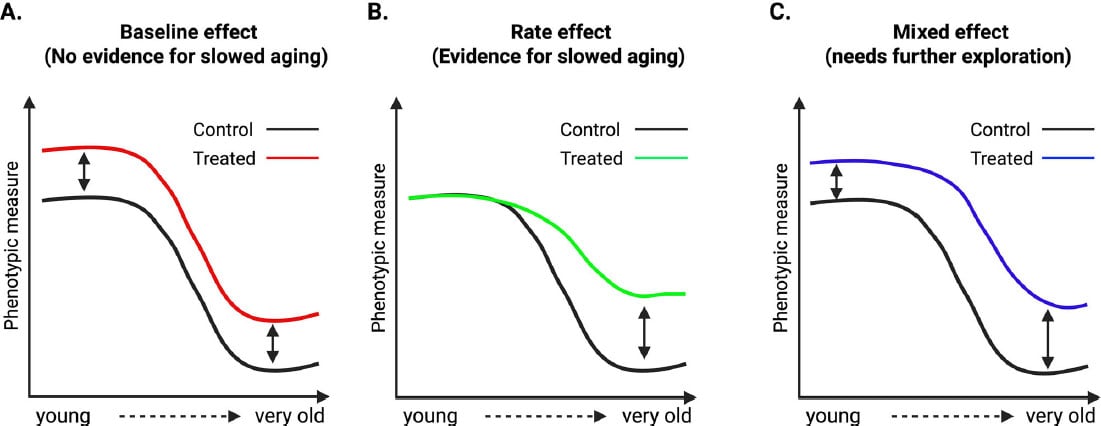
To illustrate these categories, the review highlights
studies of well-known lifespan extending interventions. These include
intermittent fasting, rapamycin, and genetic changes affecting mTOR and growth
hormone signaling. Researchers performed deep phenotyping on both young and old
animals receiving these treatments.
Although these interventions are known to extend lifespan,
many of the measured traits changed similarly in both age groups. This
indicates baseline shifts rather than slower age-related decline. In other
words, the treatments altered phenotype values but did not consistently change
the rate of aging.
Major Unanswered Questions About Aging Biology
The analysis also raises broader questions about how aging
works across the body. For instance, scientists still do not fully understand
why some tissues age faster than others or how strongly aging processes are
coordinated between organs.
Evidence shows that different tissues follow distinct aging
trajectories. These differences may arise from developmental patterns and
lifelong variation in cell turnover, metabolic activity, and exposure to
environmental stress. Researchers also continue to debate whether aging is
controlled mainly by central regulatory systems or by processes that occur
independently within individual cells.
Another challenge involves translating findings between
species. The dominant causes of death vary widely across organisms. Humans
mainly die from cardiovascular disease, mice from cancer, fish from infections,
flies from intestinal or neuromuscular failure, and worms from bacterial
infection. These differences suggest that aging may not operate as a single
universal process but instead as a complex mix of species-specific and
tissue-specific mechanisms shaped by evolution and environment.
The consequences of these findings extend beyond academic
debates. If commonly used aging markers confuse baseline physiological effects
with real changes in aging, research resources could be directed toward
treatments that improve symptoms without altering the underlying biology of
aging. The authors emphasize that geroscience aims to understand mechanisms
that drive age-related changes in physiology. Other scientific fields already
focus on regulating specific biological functions such as hormones, neural
activity, or immune responses.
Implications for Geroscience and Drug Development
For example, a therapy that improves cognitive performance
at any age may still be valuable. However, it cannot be considered a treatment
for cognitive aging unless it clearly slows the rate of cognitive decline over
time. This distinction has major implications for drug development, clinical
trial design, and expectations for patients seeking therapies that truly modify
aging.
The researchers conclude with several recommendations for
improving study design. Scientists should develop standardized panels of
age-sensitive phenotypes across multiple tissues and organ systems, covering
molecular, cellular, tissue, and whole-organism levels. Experiments should
include both young-treated and old-treated groups so that researchers can
identify whether interventions change the rate of aging or simply shift
baseline biology.
They also recommend classifying outcomes into rate effects,
baseline effects, or mixed effects rather than assuming all observed changes
reflect aging modification. Researchers should map age-sensitive trajectories
to select appropriate time points for measurement while avoiding survival bias.
Finally, claims about systemic antiaging effects should be supported by
evidence across many phenotypes and tissues rather than based on improvements
in a single outcome.
“Refining both discovery pipelines and intervention testing
frameworks will support a more mechanistic understanding of aging by enabling
researchers to distinguish between interventions that simply extend lifespan or
improve isolated age-sensitive phenotypes and those that fundamentally modify
the biological processes driving age-related decline,” the authors write.
Reference: “Beyond the hallmarks of aging: Rethinking what
aging is and how we measure it” by Maryam Keshavarz and Dan Ehninger, 2
December 2025, Genomic Psychiatry.
DOI:
10.61373/gp025i.0119
The work was supported by the ETERNITY project consortium, funded by the European Union through the Horizon Europe Marie Sklodowska-Curie Actions Doctoral Networks under grant agreement number 101072759.